In the desperate search for straw-men, AiG has plummeted to an all-time low: citing X-Men movie as an authority on mutation. Check it out for a grand ol' laugh!
Read full post
Saturday, December 31, 2005
Wednesday, December 21, 2005
Resistance is evolution; resistance is futile
It's one thing to not understand evolution, and another (though related) thing to disbelieve it. It's quite spectacular to be a public front against evolution where nearly every word you speak or write demonstrates your continued failure to understand it in the first place... like this AiG article.
In engagements with creationism, it's quite frequent to see examples of antibiotic or pesticide resistance trotted out as examples of evolution. Indeed, they are examples of the action of natural selection in changing the genetic 'make-up' of organsims. Generally, the examples are pests or microbes. So, how do creationists respond to this? The AiG article states:
After having read this paragraph, I was geared-up for how they were going to explain that this is not evolution. However, this is how I've always understood the mechanism of evolution by natural selection: variation among members of the population will cause a variation in how well individuals can reproduce. When these variations are heritable the net effect is that the advantageous variations become more widely represented in the population. Over many generations, there is a growth and eventual take-over of this heritable variation. If this process is continued and accumulated, then continued change will occur. Very simple.
The statement that this was "inherited" information and "has nothing to do with evolution" is downright laughable. I would like to know how the folks at AiG would explain evolution without inheritance.
Lest they should explain their assertions, they follow it up with this classic canard:
Ah yes - good ol' loss of information. It's never really explained what is meant by a loss of information. A loss of gene sequence? 'Overwriting' the old code? I'm not really clear on what is meant. I can only suspect this options. At the risk of putting words in their mouths, I'll focus on these options until such time as a creationist defines what they mean by 'information'.
The loss of gene sequence: Quite ofthen the case with resistance evolution. Pesticides and antimicrobials often work by 'plugging' into a key protein pathway such as to block biologically necessary reactions. A loss of a certain 'non-vital' part of a protein in a pathway may change that protein's conformation and therefore render the pathogenic agent useless.
The problem is that nobody would disagree that this type of event cannot explain how you get from, say, a self-replicating RNA to a single-celled organism. Well, not by itself (it would be naïve to think that such events never would have happened during that transformation). However, the point is irrelevant because it is wholly wrong that novel traits caused by mutations always evolve this way. Gene duplications are a common source of new adaptive capacities by microbial populations. Given that nearly all our developmental genes represent duplicate/divergent copies of the exact same genes in other animals, duplication events are precisely the type of thing we need to explain major transformations in evolutionary history.
The 'overprinting' of old information. Simply stated: this is not a loss but a changing of information. It's essential for evolution to proceed. Granted, evolution to new environments (say, those containing an anti-microbial substance) will carry antagonistic consequences for adaptations to the old environment. However, this has no effect on the acceptability of evolution. Evolution does not state that organisms are Swiss Army knives becoming infinitely large with gadgets for all environments (would sound more like something an omnipotent creator could do though). No, evolution is far from omnipotent. It is limited, imperfect, and often struggling. It can make the descendents of a fish capable of living on land, but it can't make it a universal fish/tetrapod. It loses some aquatic adaptations for life on the land. Even amphibians carry a cost of their terrestriality (such as dessication, the need to return to water). Aquatic reptiles, on the other hand, must surface to breathe. There's always a cost in evolution. The loss of adaptedness in the old environment in order to cope with a new one is not contra evolution.
The other problem is bias-laden terminology: referring to mutations as "mistakes in the genes". Indeed, they are errors in copying fidelity. Whether or not they are "mistakes" depends entirely on what their effects are. Our genomes are peppered with old genes that don't work anymore. That aside (because even creationists will challenge the 'pseudo' in 'pseudogenes'), a 'mistake' can only be interpreted in light of what effect that copying error actually has - not whether or not it changes the old sequence we're accustomed to.
So to creationists, I say that efforts to undermine evolution through the use of loaded semantics and jargon buzz-wording ('information') are futile. This is a question about data and evidence. As AiG themselves point out, observations can be interpreted different ways. However, some interpretations can be shown to be unacceptable or otherwise useless. Citing difference in interpretation does not permit using demonstrably misleading or deliberately vague interpretation.
Read full post
In engagements with creationism, it's quite frequent to see examples of antibiotic or pesticide resistance trotted out as examples of evolution. Indeed, they are examples of the action of natural selection in changing the genetic 'make-up' of organsims. Generally, the examples are pests or microbes. So, how do creationists respond to this? The AiG article states:
But in this example, it was known that this resistance to the antibiotic was already in the bacterial population right from the beginning of the experiment. In other words, some bacteria already had the information in their genes to be resistant. The bacteria had inherited information; so the resistance has nothing to do with evolution.
After having read this paragraph, I was geared-up for how they were going to explain that this is not evolution. However, this is how I've always understood the mechanism of evolution by natural selection: variation among members of the population will cause a variation in how well individuals can reproduce. When these variations are heritable the net effect is that the advantageous variations become more widely represented in the population. Over many generations, there is a growth and eventual take-over of this heritable variation. If this process is continued and accumulated, then continued change will occur. Very simple.
The statement that this was "inherited" information and "has nothing to do with evolution" is downright laughable. I would like to know how the folks at AiG would explain evolution without inheritance.
Lest they should explain their assertions, they follow it up with this classic canard:
Now, what about those experiments where some bacteria developed a resistance to substances over time due to mutations in their genes? Such mutations, which are mistakes in the genes, result from a loss of information (such as the loss of a control gene which regulates the pumping of the substance into the cell). Again, this is the opposite of evolution, which requires an increase in information if it were to occur.
Ah yes - good ol' loss of information. It's never really explained what is meant by a loss of information. A loss of gene sequence? 'Overwriting' the old code? I'm not really clear on what is meant. I can only suspect this options. At the risk of putting words in their mouths, I'll focus on these options until such time as a creationist defines what they mean by 'information'.
The loss of gene sequence: Quite ofthen the case with resistance evolution. Pesticides and antimicrobials often work by 'plugging' into a key protein pathway such as to block biologically necessary reactions. A loss of a certain 'non-vital' part of a protein in a pathway may change that protein's conformation and therefore render the pathogenic agent useless.
The problem is that nobody would disagree that this type of event cannot explain how you get from, say, a self-replicating RNA to a single-celled organism. Well, not by itself (it would be naïve to think that such events never would have happened during that transformation). However, the point is irrelevant because it is wholly wrong that novel traits caused by mutations always evolve this way. Gene duplications are a common source of new adaptive capacities by microbial populations. Given that nearly all our developmental genes represent duplicate/divergent copies of the exact same genes in other animals, duplication events are precisely the type of thing we need to explain major transformations in evolutionary history.
The 'overprinting' of old information. Simply stated: this is not a loss but a changing of information. It's essential for evolution to proceed. Granted, evolution to new environments (say, those containing an anti-microbial substance) will carry antagonistic consequences for adaptations to the old environment. However, this has no effect on the acceptability of evolution. Evolution does not state that organisms are Swiss Army knives becoming infinitely large with gadgets for all environments (would sound more like something an omnipotent creator could do though). No, evolution is far from omnipotent. It is limited, imperfect, and often struggling. It can make the descendents of a fish capable of living on land, but it can't make it a universal fish/tetrapod. It loses some aquatic adaptations for life on the land. Even amphibians carry a cost of their terrestriality (such as dessication, the need to return to water). Aquatic reptiles, on the other hand, must surface to breathe. There's always a cost in evolution. The loss of adaptedness in the old environment in order to cope with a new one is not contra evolution.
The other problem is bias-laden terminology: referring to mutations as "mistakes in the genes". Indeed, they are errors in copying fidelity. Whether or not they are "mistakes" depends entirely on what their effects are. Our genomes are peppered with old genes that don't work anymore. That aside (because even creationists will challenge the 'pseudo' in 'pseudogenes'), a 'mistake' can only be interpreted in light of what effect that copying error actually has - not whether or not it changes the old sequence we're accustomed to.
So to creationists, I say that efforts to undermine evolution through the use of loaded semantics and jargon buzz-wording ('information') are futile. This is a question about data and evidence. As AiG themselves point out, observations can be interpreted different ways. However, some interpretations can be shown to be unacceptable or otherwise useless. Citing difference in interpretation does not permit using demonstrably misleading or deliberately vague interpretation.
Read full post
Victory for reason in Dover
It doesn't get any better than this. A ruling against the ID pundits was forseeable, but the frank and downright scathing tone of Judge Jones ruling was the icing on the cake. It quite possibly stands as the single most indicting document against ID ever.
Judge Jones did not shy away from pointing out liars and various dishonest and underhanded measures used by the IDists in order to advance their case. His statement bears numerous references to the over religiosity of ID.
Need I say more? I think this event speaks volumes for itself. I think I'll raise a glass tonight.
Cheers!
Read full post
Judge Jones did not shy away from pointing out liars and various dishonest and underhanded measures used by the IDists in order to advance their case. His statement bears numerous references to the over religiosity of ID.
Need I say more? I think this event speaks volumes for itself. I think I'll raise a glass tonight.
Cheers!
Read full post
Monday, December 12, 2005
Species is as species does... Part II - What teeth will tell
Another question that comes up is more specific, but is also an example drawn from dogs. It's frequently charged that if a palaeontologist had found the skeleton of a thylacine (or Tasmanian 'wolf' or Tasmanian 'tiger'), they would probably classify it (incorrectly) as a dog, rather than as a marsupial. The reason, we are told, is because the skeleton of a thylacine "looks like" a dog's, more than it does that of a marsupial such as a kangaroo or koala.
Here's a wolf skull. For our purposes, entirely like that of a typical domestic dog.

Now here's the skull of a thylacine

Upon first inspection, it's hard to see how a palaeontologist could possibly distinguish the two skulls. The reality is that it's very, very easy. Turn the skull over and look at the teeth. Here's dental arcade of a thylacine:

Note that there are four molars, the last three being nearly identical except in size. Now look at the tooth arcade of a dog:
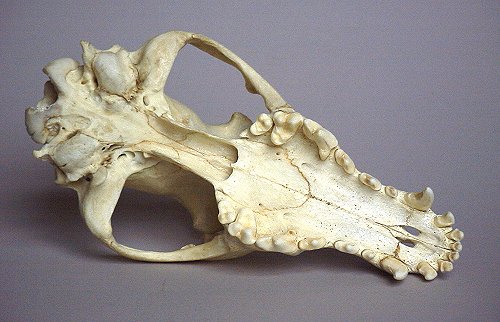
One of the defining features of modern placental mammals is that they never have more than three molars. This already separates the thylacine skull not only from dogs, but all placentals. In this case, the dog only has two molars and a single premolar modified into what is called a 'carnassial' - a shear-like tooth for cutting meat. Note also that the thylacine skull has a pair of palatal fenestrae: window-like openings on the bottom of the skull, a character found in marsupials such as wombat skull. The dog skull clearly does not. (Don't confuse it with the passages seen behind the dog's palate - this is the choana, or internal nostril. The thylacine has this feature in addition to the palatal fenestrae, but it's not seen in the picture). These actually amount to only a few of the numerous differences in the skull that distinguish placental mammals from marsupials.
The fact that systemtatists and palaeontologists can identify marsupials from their bones is evidenced by the sabre-toothed marsupials. These forms look very similar to the familiar sabre-tooth cats. Despite only being known from fossils, they have been distinguished by palaeontologists as marsupial and placental carnivoran, respectively. This is because there are numerous aspects of the skull, such as dental characters discussed above, that can be used to unambiguously distinguish them. In addition to skull and dental characters, marsupials bear a prepubic bone (ofthen called the "marsupial bone") that projects forward from the front of the hips. It appears to be related to the pouch that marsupials use to carry their young. When entire skeletons are available, it serves as nother character to distinguish these two groups.
Here we see that not only would palaeontologists know how to separate the thylacine from dogs on the basis of bones alone, but that they routinely do distinguish marsupials from placentals in the fossil record. The evolutionary history of organisms is long and complex. Even if two organisms happen to take on very similar general appearance, the traces of their ancestry are often still documented by other parts of their anatomy. Palaeontologists and systematists search for these traces which are held in the shared unique characters held between members that share common ancestors - rather than relying on overt general similarities.
Just as in other sciences, common sense often betrays reality in systematics. This is not to say that reality never betrays systematics, either - it certianly can. Indeed convergence and incompleteness can frequently play havoc with the reconstruction of phylogeny (evolutionary relationships). However, sticking things together by overt similarity is not how things are done. Systematists look for nested sets of these shared unique similarities which, as we have seen with dogs and now marsupial vs. placental mammals, that are most useful in uncovering the true nature of evolutionary relationships.
Read full post
Here's a wolf skull. For our purposes, entirely like that of a typical domestic dog.
Now here's the skull of a thylacine
Upon first inspection, it's hard to see how a palaeontologist could possibly distinguish the two skulls. The reality is that it's very, very easy. Turn the skull over and look at the teeth. Here's dental arcade of a thylacine:
Note that there are four molars, the last three being nearly identical except in size. Now look at the tooth arcade of a dog:
One of the defining features of modern placental mammals is that they never have more than three molars. This already separates the thylacine skull not only from dogs, but all placentals. In this case, the dog only has two molars and a single premolar modified into what is called a 'carnassial' - a shear-like tooth for cutting meat. Note also that the thylacine skull has a pair of palatal fenestrae: window-like openings on the bottom of the skull, a character found in marsupials such as wombat skull. The dog skull clearly does not. (Don't confuse it with the passages seen behind the dog's palate - this is the choana, or internal nostril. The thylacine has this feature in addition to the palatal fenestrae, but it's not seen in the picture). These actually amount to only a few of the numerous differences in the skull that distinguish placental mammals from marsupials.
{kind=link}
The fact that systemtatists and palaeontologists can identify marsupials from their bones is evidenced by the sabre-toothed marsupials. These forms look very similar to the familiar sabre-tooth cats. Despite only being known from fossils, they have been distinguished by palaeontologists as marsupial and placental carnivoran, respectively. This is because there are numerous aspects of the skull, such as dental characters discussed above, that can be used to unambiguously distinguish them. In addition to skull and dental characters, marsupials bear a prepubic bone (ofthen called the "marsupial bone") that projects forward from the front of the hips. It appears to be related to the pouch that marsupials use to carry their young. When entire skeletons are available, it serves as nother character to distinguish these two groups.
{kind=link}
{kind=link}
Here we see that not only would palaeontologists know how to separate the thylacine from dogs on the basis of bones alone, but that they routinely do distinguish marsupials from placentals in the fossil record. The evolutionary history of organisms is long and complex. Even if two organisms happen to take on very similar general appearance, the traces of their ancestry are often still documented by other parts of their anatomy. Palaeontologists and systematists search for these traces which are held in the shared unique characters held between members that share common ancestors - rather than relying on overt general similarities.
Just as in other sciences, common sense often betrays reality in systematics. This is not to say that reality never betrays systematics, either - it certianly can. Indeed convergence and incompleteness can frequently play havoc with the reconstruction of phylogeny (evolutionary relationships). However, sticking things together by overt similarity is not how things are done. Systematists look for nested sets of these shared unique similarities which, as we have seen with dogs and now marsupial vs. placental mammals, that are most useful in uncovering the true nature of evolutionary relationships.
Read full post
'Species' - Part II
Here's something annoying about using Blogger: it publishes your post in queue according to the day you started the draft not on the day you chose to publish. As a result, here's a link to Part II of 'Species is as species does..." which now appears a few posts down (as opposed to in the place of this... grrr!
Edit (15/12/05): Well, after a few days and the help of Coturnix, my eyes suddenly gained the ability to see the "Change Time & Date" feature! My oh my. I have to stop writing these late at night!
Read full post
Edit (15/12/05): Well, after a few days and the help of Coturnix, my eyes suddenly gained the ability to see the "Change Time & Date" feature! My oh my. I have to stop writing these late at night!
Read full post
Saturday, December 10, 2005
Infidel Guy appearance
The show went really well and lasted two hours! I was able to cover a very broad range of topics and feel quite satisfied. I've been getting a lot of good feedback, so it sounds like people were both enjoying it and learning something. If you're a member over at Infidel Guy you can download a recording of the show.
Read full post
Read full post
Wednesday, December 07, 2005
Internet radio appearance: Live With the Infidel Guy
For those of you who don't know, I'll be appearing this Friday at 8PM Eastern (December 9, 2005) on the Infidel Guy internet radio show with Reggie Finley. I'll be talking about evolution and the fossil record and taking up a few of those really popular creationist canards. I believe the show takes callers if you're interested in asking questions.
Edit: I'd like to welcome all the new traffic from the Infidel Guy site! Thanks for stopping by, I hope you find something of interest here!
Read full post
Edit: I'd like to welcome all the new traffic from the Infidel Guy site! Thanks for stopping by, I hope you find something of interest here!
Read full post
Species is as species does... Part I - Variation vs. Speciation
People often ask me about species in the fossil record, and they are often posing some good questions. This will be the first of a three-part series of entries that I am going to do on this topic.
"How do we know that two fossils - often very similar - are different species?" The especially inquisitive person will often point out an array of different dog skulls and likens it to the similar array of hominin skulls. "How do we know that these alleged hominin species can't interbreed?" The short answer is that we don't know. The difficult part is explaining why it doesn't matter all that much.
Much of the problem seems to stem from an attachment to the Biological Species Concept (BSC) as canonical truth and that knowledge of breeding compatibility has everything to do with reconstructing phylogeny. The BSC states that a species is a population of organisms that can interbreed and produce fertile offspring. It's easy to see why it causes alarm, because it obviously can't be applied to fossils. It's also invalid when applied to asexual organisms (of which there are a great, great many).
In a somewhat concerned tone, this person might also state their belief that, were the different breeds of dogs to be extinct and their skulls discovered by palaeontologists today, we might well classify them as different species. Oh the horror! They would be so dismally wrong and never even know it! Wouldn't this cause dramatic consequences for our phylogenies of animals? The answer is that it's unlikely because of the way phylogenies are reconstructed (as opposed to the way people think they're constructed).
Palaeontologists employ a modern method of systematic biology called cladistics. Cladistics ignores characters unique to individual species because these don't tell us anything about relationships. For instance, if we're trying to figure out how a cow, a trout, and a human are related, the fact that a human has an opposable thumb tells us nothing about how it is related to the cow or the trout, as neither have it. Similarly, the fact that all three have a backbone is useless (except in setting the three apart from invertebrates, but we haven't stipulated that). Instead, cladistics focuses on character that are unique and shared between broader collections of species (i.e. four limbs with digits are uniqu and shared between humans and cows, to the exclusion of the trout).
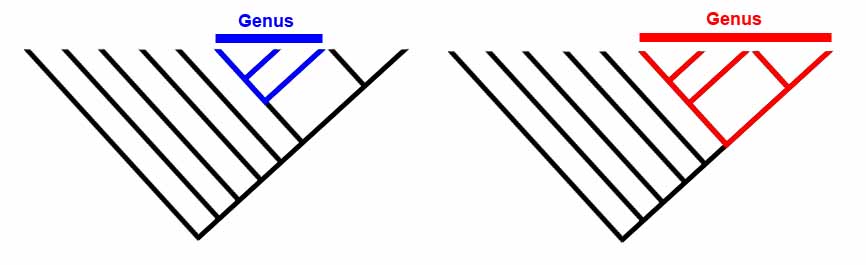
But first, let's consider how the assumption of species might affect our methods of determining relationships in the first place. Let's imagine that dogs have been extinct for several million years and that palaeontologists have discoverd a bunch of different dog skulls. They've named each distinct type of skull a new species. Suppose that the result of their cladistic analysis looks like this:
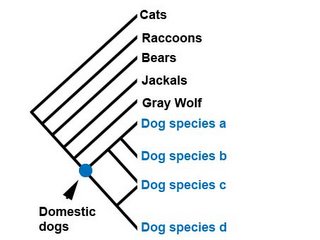
Systematists working on extant (i.e. not extinct) animals also use the very same techniques that palaeontologists use (but will employ molecular data, not just morphological data wherever they can). Assuming that the phylogenies are the correct ones, they should get a result that looks like this:
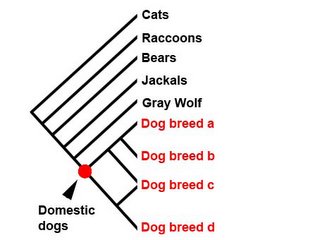
The interrelationships should be identical. The differences are only in the very tips of the branches (breeds vs. species). The assumption of 'species' or 'breeds' is trivial, because the results are the same. The question, however, is would the same result be obtained were it not already known that dogs are interfertile?
The modern complement of dog breeds are (barring some mechanical difficulties) interfertile. They are, apparently, also interfertile with wolves. If we look at an array of dogs, we can see that a key character unites all of the domestic ones: the raised frontal (or 'forehead'):

This clearly sets them apart from foxes (top) and coyotes (bottom):


(by the way, all these great skull pictures are from this fantastic site)
What should be immediately obvious is that, despite the differences between the breeds of domestic dogs, their recent common ancestry with each other is general character that they share in common to the exclusion of other canids and carnivores. This is why modern systematics focuses mostly on shared similarities, rather than on separating them by differences. Because of this fact, domestic dogs (as we know them) would probably have been classified as a single genus, even on the basis of their morphology. Perhaps they would have been classified as distinct species, but as a total group they would have been recognized as distinct from all other canids. Historically speaking, this is identical to the true phylogeny. This is not to say that we'll always have the right answer. Rather, it is to say that by being sufficiently prudent about our classification, we should still be able to uncover the broad-scale patterns of evolution, at the very least.
For palaeontologists, it will never be possible to determine interbreeding relationships. But as long as we don't take the BSC as canon (and we shouldn't), then there is no problem. Naming a new species in palaeontology simply reflects being able to point to a distinctive morphology - preferably if we can show that populations of fossil animals can similarly be categorized. If we're concerned about whether or not two different fossil species (or perhaps even genera!) are interfertile, we are hopeless. However, we should still be able to uncover relative common ancestry. In the diagrams below, we can see that the trees (evolutionary relationships) are identical. However, the taxonomy is quite different.
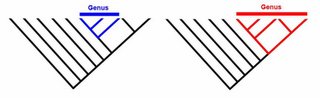
So a species is as species does: for the purposes of palaeontology, it identifies a range of morphological variation. Since species/breeds are always nested the same way within genera (more inclusive groups of variation), it doesn't really matter if we call them species, breeds, or varieties in fossil animals. As long as they're within the same genus (and don't take the BSC to heart), then we're saying exactly the same thing about the course of evolution. Indeed, there is no question that all the different skulls of the different dog breeds would be classified in the same genus, along with wolves based on morphology.
I'll follow this up in Part II of this post, where I will discuss another nagging question: the classification of dogs and the Tazmanian 'wolf'. If we didn't know it from living specimens (though, thanks to man, it is now extinct), would it not have been classified as a dog?
Read full post
"How do we know that two fossils - often very similar - are different species?" The especially inquisitive person will often point out an array of different dog skulls and likens it to the similar array of hominin skulls. "How do we know that these alleged hominin species can't interbreed?" The short answer is that we don't know. The difficult part is explaining why it doesn't matter all that much.
Much of the problem seems to stem from an attachment to the Biological Species Concept (BSC) as canonical truth and that knowledge of breeding compatibility has everything to do with reconstructing phylogeny. The BSC states that a species is a population of organisms that can interbreed and produce fertile offspring. It's easy to see why it causes alarm, because it obviously can't be applied to fossils. It's also invalid when applied to asexual organisms (of which there are a great, great many).
In a somewhat concerned tone, this person might also state their belief that, were the different breeds of dogs to be extinct and their skulls discovered by palaeontologists today, we might well classify them as different species. Oh the horror! They would be so dismally wrong and never even know it! Wouldn't this cause dramatic consequences for our phylogenies of animals? The answer is that it's unlikely because of the way phylogenies are reconstructed (as opposed to the way people think they're constructed).
Palaeontologists employ a modern method of systematic biology called cladistics. Cladistics ignores characters unique to individual species because these don't tell us anything about relationships. For instance, if we're trying to figure out how a cow, a trout, and a human are related, the fact that a human has an opposable thumb tells us nothing about how it is related to the cow or the trout, as neither have it. Similarly, the fact that all three have a backbone is useless (except in setting the three apart from invertebrates, but we haven't stipulated that). Instead, cladistics focuses on character that are unique and shared between broader collections of species (i.e. four limbs with digits are uniqu and shared between humans and cows, to the exclusion of the trout).
But first, let's consider how the assumption of species might affect our methods of determining relationships in the first place. Let's imagine that dogs have been extinct for several million years and that palaeontologists have discoverd a bunch of different dog skulls. They've named each distinct type of skull a new species. Suppose that the result of their cladistic analysis looks like this:

Systematists working on extant (i.e. not extinct) animals also use the very same techniques that palaeontologists use (but will employ molecular data, not just morphological data wherever they can). Assuming that the phylogenies are the correct ones, they should get a result that looks like this:

The interrelationships should be identical. The differences are only in the very tips of the branches (breeds vs. species). The assumption of 'species' or 'breeds' is trivial, because the results are the same. The question, however, is would the same result be obtained were it not already known that dogs are interfertile?
The modern complement of dog breeds are (barring some mechanical difficulties) interfertile. They are, apparently, also interfertile with wolves. If we look at an array of dogs, we can see that a key character unites all of the domestic ones: the raised frontal (or 'forehead'):

This clearly sets them apart from foxes (top) and coyotes (bottom):
(by the way, all these great skull pictures are from this fantastic site)
What should be immediately obvious is that, despite the differences between the breeds of domestic dogs, their recent common ancestry with each other is general character that they share in common to the exclusion of other canids and carnivores. This is why modern systematics focuses mostly on shared similarities, rather than on separating them by differences. Because of this fact, domestic dogs (as we know them) would probably have been classified as a single genus, even on the basis of their morphology. Perhaps they would have been classified as distinct species, but as a total group they would have been recognized as distinct from all other canids. Historically speaking, this is identical to the true phylogeny. This is not to say that we'll always have the right answer. Rather, it is to say that by being sufficiently prudent about our classification, we should still be able to uncover the broad-scale patterns of evolution, at the very least.
For palaeontologists, it will never be possible to determine interbreeding relationships. But as long as we don't take the BSC as canon (and we shouldn't), then there is no problem. Naming a new species in palaeontology simply reflects being able to point to a distinctive morphology - preferably if we can show that populations of fossil animals can similarly be categorized. If we're concerned about whether or not two different fossil species (or perhaps even genera!) are interfertile, we are hopeless. However, we should still be able to uncover relative common ancestry. In the diagrams below, we can see that the trees (evolutionary relationships) are identical. However, the taxonomy is quite different.
So a species is as species does: for the purposes of palaeontology, it identifies a range of morphological variation. Since species/breeds are always nested the same way within genera (more inclusive groups of variation), it doesn't really matter if we call them species, breeds, or varieties in fossil animals. As long as they're within the same genus (and don't take the BSC to heart), then we're saying exactly the same thing about the course of evolution. Indeed, there is no question that all the different skulls of the different dog breeds would be classified in the same genus, along with wolves based on morphology.
I'll follow this up in Part II of this post, where I will discuss another nagging question: the classification of dogs and the Tazmanian 'wolf'. If we didn't know it from living specimens (though, thanks to man, it is now extinct), would it not have been classified as a dog?
Read full post
Saturday, December 03, 2005
More rhizodonts
My first paper was published two weeks ago in the Canadian Journal of Earth Sciences (Canadian visitors have free online access), so I figure I should put up a notice on here. The work describes a new genus of rhizodontid fish from the Early Carboniferous of Nova Scotia. It was the subject of my undergraduate thesis.
Read full post
Read full post
Saturday, November 12, 2005
Peptide Antibiotics: Boot Camp for Bacteria?
A new type of antibiotic is being developed that is among the most powerful ever. Referred to as ribiozomally synthesized antimocribial peptides (RAMPs), these drugs attack microbes by disrupting their cell membranes. The drug effective spills the guts of bacteria by breaking them open. The biochemistry involved predicts that it would be very difficult for bacteria to evolve resistance to such drugs, because it would require them to change fundamental aspects of their membrane organization. Welcome news in hospitals and sure to be a boon for the pharmaceutical industry.
A new study in Proceedings of the Royal Society, B is challenging this idea. Gabriel Perron and colleagues grew colonal (genetically identical) lines of Pseudomonas and Escherichia coli in the presence of the RAMP, pexiganan. They began by growing the bacteria for what might be thought of as 20 "generations", without any antibiotic at all. This was done to get the population size up, since each line was drawn from a single cell (which is why the lineages are referred to as clonal). Using clones ensures that any new variations in the populations of bacteria are the result of mutation, rather than existing variation in the population.
Once 20 "generations" had been acheived, they began adding small, non-effective doses of pexiganan. Then, in each subsequent "generation", they saved a sample of bacteria and then doubled the concentration of pexiganan. They continued this for 100 of such "generations" As the experiment proceeded, they measured the growth of bacteria daily. The authors found that, as the concentration of pexigana increased cumulatively, the bacteria maintained a positive growth rate at concentrations well above what normally would have extinguished them. The bacteria had evolved resistance.
Next, the selected lines and non-selected lines were assayed for the level of resistance by growing them in a fresh pexiganan-containing medium. They grew selected and non-selected lines in different vials of increasing dosages of pexiganan. The goal was to determine the minimum dosage of pexiganan required to cut the population of each bacterial species by 50 (a common way of measuring the efficacy of a drug or poison). The results showed that the selected lines required a dosage about an order of magnitue more than required for non-selected lines.
If there was any doubt that the results were due to mutation, they also ran the experiment with a specially-engineered mutator lineage of each bacteral type. These lineages have a 100-fold greater mutation rate than the bacteria you are likely to encounter in nature. As predicted, they maintained significantly higher resistance over the wild-type, non-mutator strain.
Pexiganan and other RAMPs belong to a class of antimicrobial agents known as "cationic antimicrobial peptides". Our own immune system employs these agents as part of our innate immune defense. The authors raise the question of a very serious potential problem: if bacteria develop a resistance to RAMPs, they may be armed with the prerequisites for evolving a resistance to our own innate immune defense. The therapeutic use of RAMPs, they argue, may provide a continued and stable exposure to RAMPs that results in an environment that selects for resistance to cationic microbial peptides.
Perron et al. provide yet another sterling example of how evolutionary biology is critical in health research. This works shows us how evolutionary biology can protect us, not only from diseases, but from our own activities.
Read full post
A new study in Proceedings of the Royal Society, B is challenging this idea. Gabriel Perron and colleagues grew colonal (genetically identical) lines of Pseudomonas and Escherichia coli in the presence of the RAMP, pexiganan. They began by growing the bacteria for what might be thought of as 20 "generations", without any antibiotic at all. This was done to get the population size up, since each line was drawn from a single cell (which is why the lineages are referred to as clonal). Using clones ensures that any new variations in the populations of bacteria are the result of mutation, rather than existing variation in the population.
Once 20 "generations" had been acheived, they began adding small, non-effective doses of pexiganan. Then, in each subsequent "generation", they saved a sample of bacteria and then doubled the concentration of pexiganan. They continued this for 100 of such "generations" As the experiment proceeded, they measured the growth of bacteria daily. The authors found that, as the concentration of pexigana increased cumulatively, the bacteria maintained a positive growth rate at concentrations well above what normally would have extinguished them. The bacteria had evolved resistance.
Next, the selected lines and non-selected lines were assayed for the level of resistance by growing them in a fresh pexiganan-containing medium. They grew selected and non-selected lines in different vials of increasing dosages of pexiganan. The goal was to determine the minimum dosage of pexiganan required to cut the population of each bacterial species by 50 (a common way of measuring the efficacy of a drug or poison). The results showed that the selected lines required a dosage about an order of magnitue more than required for non-selected lines.
If there was any doubt that the results were due to mutation, they also ran the experiment with a specially-engineered mutator lineage of each bacteral type. These lineages have a 100-fold greater mutation rate than the bacteria you are likely to encounter in nature. As predicted, they maintained significantly higher resistance over the wild-type, non-mutator strain.
Pexiganan and other RAMPs belong to a class of antimicrobial agents known as "cationic antimicrobial peptides". Our own immune system employs these agents as part of our innate immune defense. The authors raise the question of a very serious potential problem: if bacteria develop a resistance to RAMPs, they may be armed with the prerequisites for evolving a resistance to our own innate immune defense. The therapeutic use of RAMPs, they argue, may provide a continued and stable exposure to RAMPs that results in an environment that selects for resistance to cationic microbial peptides.
Perron et al. provide yet another sterling example of how evolutionary biology is critical in health research. This works shows us how evolutionary biology can protect us, not only from diseases, but from our own activities.
Read full post
Monday, October 10, 2005
Phony feathered dinosaurs?
According to a recent article from the Journal of Morphology (subscription required), the integumentary filaments of those famous feathered dinosaurs are nto feathers at all. According to Alan Feduccian and colleagues, the filaments observed are evidence of decaying collagen. Fair enough. This would be very interesting to know, and would be a welcome challenge. The problem is that since Feduccia and company have been embarrassed countless times, they will do most anything to try to save face. This massive, 47-page article in J. Morph. stands as a testiment to their zombie-like resiliance.
While the integumentary filaments of the dinosaurs from eastern China may, in some instances, represent frayed collagen fibers from the sub-epidermal tissues there are a number of patterns which remain to be explained. Chief among them: why don't the countless crocodiles, lizards, and turtles from these sites show similar filaments?. Why is it that only the birds and the dinosaurs from the Jehol biota happen to have these integumentary filaments? Do crocodiles, lizards, and turtles not have these? That would be really interesting since Feduccia et al. referred to specimens of each of these groups for their studies. They provided no comparative evidence from other reptiles from this site that would corroborrate their hypothesis.
Especially interesting is the following statement. Because Feduccia et al. have already decided that birds could not possibly come from dinosaurs, they choose to dispute the phylogenetic position of feathered dinosaurs such as Caudipteryx with this kind of hand-waving:
Well, how convenient. Little do they realize, it doesn't matter if these animals are secondarily flightless birds. The question of explaining the enormous number of characters they share in common with birds remains the important evolutionary question. They have simply chosen to deny the overwhelming character data in favour of their pre-conceived notion that birds could not have descended from dinosaurs.
The absolute icing on the cake is this tacit appeal by Feduccia to creationist reasoning as support for his ideas, from Science Daily:
This is not just pure bollocks, it's blatantly irresponsible. Thanks a lot Feduccia! Thanks very much for that. While you're at it, why don't you cite the Grand Canyon as evidence against modern geology! Good job!
More on this tomorrow. It's late here.
Read full post
While the integumentary filaments of the dinosaurs from eastern China may, in some instances, represent frayed collagen fibers from the sub-epidermal tissues there are a number of patterns which remain to be explained. Chief among them: why don't the countless crocodiles, lizards, and turtles from these sites show similar filaments?. Why is it that only the birds and the dinosaurs from the Jehol biota happen to have these integumentary filaments? Do crocodiles, lizards, and turtles not have these? That would be really interesting since Feduccia et al. referred to specimens of each of these groups for their studies. They provided no comparative evidence from other reptiles from this site that would corroborrate their hypothesis.
Especially interesting is the following statement. Because Feduccia et al. have already decided that birds could not possibly come from dinosaurs, they choose to dispute the phylogenetic position of feathered dinosaurs such as Caudipteryx with this kind of hand-waving:
That phylogenetics has become an assumptionladen
field is best illustrated by the insistence that
the avian wing of Caudipteryx, with its intricate
detailed flight anatomy and avian arrangement of
primary and secondary feathers on the hand and
arm (Fig. 26), evolved in a context other than flight
Well, how convenient. Little do they realize, it doesn't matter if these animals are secondarily flightless birds. The question of explaining the enormous number of characters they share in common with birds remains the important evolutionary question. They have simply chosen to deny the overwhelming character data in favour of their pre-conceived notion that birds could not have descended from dinosaurs.
The absolute icing on the cake is this tacit appeal by Feduccia to creationist reasoning as support for his ideas, from Science Daily:
The theory that birds are the equivalent of living dinosaurs and that dinosaurs were feathered is so full of holes that the creationists have jumped all over it, using the evolutionary nonsense of ‘dinosaurian science’ as evidence against the theory of evolution
This is not just pure bollocks, it's blatantly irresponsible. Thanks a lot Feduccia! Thanks very much for that. While you're at it, why don't you cite the Grand Canyon as evidence against modern geology! Good job!
More on this tomorrow. It's late here.
Read full post
Wednesday, September 21, 2005
Understanding Evolution
Berkely has launched their new and improved Understanding Evolution page. It's colourful, inviting, uses a tasteful number of flash animations, and still looks professional. It's very easy to navigate, but they really ought to put the beef near the top. A guided tour through evolution is what people need to see.
Unfortunately, there are a couple problems I see with this site. It spends too much time presenting the formulation of evolution. That is, it presents too much about how "we think" evolution works. I agree this is important, because creationists have done such a fine job of creating straw men out of age-old public misconceptions. Unfortunately, much of what I see will re-inforce creationist prejudices that we are just spoon-feeding them an "alternate truth" to the one they were initially fed. Which "truth" do you think they'll go for?
The presentation of homology as evidence for evolution is, I'm sorry to say, bad, bad, bad. I'm going to give some credit here to creationists in order to point out why this is bad. First, the Berkely site starts with the classic example of tetrapod limbs. While I agree it's a very ostensible example, one must remember that an evolution-weary mind is approaching this with doubts and reservations. Creationists already have a response to this and this new presentation makes no pre-emptive strikes..
Here's the problem, the skeptical student (doubful, but interested in knowing more about evolution) is faced with a dichotomy:
similar structures are caused by a common ancestor
-or-
similar structures are caused by a common designer
Why is one of these statements better than the other?
To most biologists, it's clear why the first is a better explanation. It actually offers a reason to expect the pattern, whereas the second relies on some statement about the capriciousness of the deity. To the general (and perhaps somewhat skeptical) reader, this is not so clear. In fact, both of the above statements could be viewed as begging the question (i.e. "circular reasoning").
The important issue is not simply one of similarity, but rather of similarity distributions. To which animals do these sorts of limbs belong? Why don't flies or starfish have limbs with these bones in them? Such limbs always belong to special subset of vertebrate animals. The emerging picture is one of a nested hierarchy which really is the key issue. Since nested hierarchies are a mathematically necessary consequence of any branching process (i.e. common ancestry) we have a priori reasons to expect such a pattern to emerge. This is the actual theoretical framework that is the other half of the agument. The reasons for expecting this.
In the context of a nested hierarchy, the distribution of limb skeletons is precisely and explicity predicted by a theory of common ancestry. What students are never presented with first is a picture of nested hierarchies. They're thus not presented with a picture of the evidence for common ancestry. Instead, they're first presented with a picture of a tree (the inference) and a bunch of characters mapped on (the evidence). To a naïve and dubious young student, this looks as tenuous as any crackpot seminar or tract booklet. It's easily charicatured as selective data usage and has become a favourite of creationist attacks on evolution education. Show the kids the evidence first, then show them how the conclusions came from that evidence.
If you're going to present limb homologies to students, then you have to have other evidence to re-inforce your conclusion. Otherwise, you're just going to get the "common designer" crap in return. The theory of evolution is far too elegant to be in competition with such an illogical argument.
#
Read full post
Tuesday, September 20, 2005
Little Piggies of Urodeles
A recent set of exchanges between myself and a friend brought me upon this Answers in Genesis article. In it, Jonathan Sarfati (an acid-tongued spokesperson for AiG) claims that the digit development of frogs is completely different from that of all other tetrapods. In it, he claims, frogs grow their digits from developing buds. PZ Myers responded to Sarfati's claims about bird/dinosaur digit homology and amphibian digit development.
A minor correction is worth publishing. In the article Myers states:
Despite the fact that I think most of Myers's article is a wonderful vulgarization of limb development, this last part is an overstatement.
It is true that many aspects of limb and digit development in tetrapods is remarkably well conserved throughout tetrapods, but it is not true that all tetrapods have an AER, it is also not true that they all use apoptosis to create the interdigital spaces. All salamanders that have so far been studied grow their digits from buds and lack an AER, much like Sarfati explained for frogs (only Sarfati attributes it to the wrong animal). Apoptosis has only been detected in the developing digits of one salamander Desmognathus and its role in limb development is not clear (Franssen et al. 2005). It is also the case that salamanders begin digit development with digit 2, which is also in stark contrast with the majority of other tetrapods which develop digit 4 first. This is an interesting and perplexing problem for developmental biologists.
So does this mean that Sarfati is right? Does this mean that the limbs of salamanders cannot be homolgous to those of other tetrapods? The problem comes from this statement by Sarfati which exemplifies a rather deep ignorance of developmental biology (and I'm not even an expert!):
This is both overly simplistic and fallacious, since we already know that most of the "genetic toolkit" for animal development is highly conserved. The fact is, the same genes are used in nearly all tetrapods. In fact, vast numbers of the same genes are involved in similar processes in all animals. Some genes have been duplicated and modified to produce different functions -- which is precisely how animal form evolves!
As Myers points out beautifully, and what is the most important point here, is that the patterning mechanisms are conserved. We know that developing tetrapod limbs use a special hierarchical pattern of Hox genes that lay out the "construction zones" in a developing embryo. We see remarkable conservation in both the Hox genes and the placement of special region known as the zone of polarizing activity (ZPA) which plays a role in setting up anterior-posterior polarity of the developing limb.
The emerging picture of salamander limb development is that there are a striking number of "mechanical" differences in the way the construction takes place. The order of digit development and many of the processes that make the digit protrusions are different. However, what doesn't appear to have changed, is precisely what we would expect if the digits are homologous: the genes that control the identity of the digits.
Animal development doesn't work in the same way as humans construct buildings. It's not as logical as we would think it is. In fact, it's rather perplexing and enigmatic and often proceeds along seemingly very illogical courses. The process of making a digit (as in, a protrusion of the distal part of a limb) is under different controls from the process that says what those digits are (the thumb vs. forefinger, etc.). Sarfati's oversimplification is academically irresponsible. It is flat-out wrong making the entire thesis of his argument baseless.
Franssen, R.A., Marks, S., Wake, D. and Shubin, N. 2005. Limb chondrogenesis of the seepage salamander (Desmognathus aenus(Amphibia: Plethodontidae). Journal of Morphology 265:87-101
Read full post
A minor correction is worth publishing. In the article Myers states:
We’ve got a pretty good handle on the outline of limb development in multiple tetrapod lineages now, and they all use the same tools. Contrary to Sarfati’s implication, they all have apical ectodermal ridges (with some rare exceptions in a few highly derived, direct-developing frogs) and zones of polarizing activity, they all use the same set of molecules, including FGF-4 and FGF-8 and the same Hox genes and retinoic acid and BMPs. If there’s one thing we know, it’s that limb development is dazzlingly well conserved.
Despite the fact that I think most of Myers's article is a wonderful vulgarization of limb development, this last part is an overstatement.
It is true that many aspects of limb and digit development in tetrapods is remarkably well conserved throughout tetrapods, but it is not true that all tetrapods have an AER, it is also not true that they all use apoptosis to create the interdigital spaces. All salamanders that have so far been studied grow their digits from buds and lack an AER, much like Sarfati explained for frogs (only Sarfati attributes it to the wrong animal). Apoptosis has only been detected in the developing digits of one salamander Desmognathus and its role in limb development is not clear (Franssen et al. 2005). It is also the case that salamanders begin digit development with digit 2, which is also in stark contrast with the majority of other tetrapods which develop digit 4 first. This is an interesting and perplexing problem for developmental biologists.
So does this mean that Sarfati is right? Does this mean that the limbs of salamanders cannot be homolgous to those of other tetrapods? The problem comes from this statement by Sarfati which exemplifies a rather deep ignorance of developmental biology (and I'm not even an expert!):
If the birds evolved from dinosaurs, then one would expect common genes. These in turn would code for a common development in the embryo.
This is both overly simplistic and fallacious, since we already know that most of the "genetic toolkit" for animal development is highly conserved. The fact is, the same genes are used in nearly all tetrapods. In fact, vast numbers of the same genes are involved in similar processes in all animals. Some genes have been duplicated and modified to produce different functions -- which is precisely how animal form evolves!
As Myers points out beautifully, and what is the most important point here, is that the patterning mechanisms are conserved. We know that developing tetrapod limbs use a special hierarchical pattern of Hox genes that lay out the "construction zones" in a developing embryo. We see remarkable conservation in both the Hox genes and the placement of special region known as the zone of polarizing activity (ZPA) which plays a role in setting up anterior-posterior polarity of the developing limb.
The emerging picture of salamander limb development is that there are a striking number of "mechanical" differences in the way the construction takes place. The order of digit development and many of the processes that make the digit protrusions are different. However, what doesn't appear to have changed, is precisely what we would expect if the digits are homologous: the genes that control the identity of the digits.
Animal development doesn't work in the same way as humans construct buildings. It's not as logical as we would think it is. In fact, it's rather perplexing and enigmatic and often proceeds along seemingly very illogical courses. The process of making a digit (as in, a protrusion of the distal part of a limb) is under different controls from the process that says what those digits are (the thumb vs. forefinger, etc.). Sarfati's oversimplification is academically irresponsible. It is flat-out wrong making the entire thesis of his argument baseless.
Franssen, R.A., Marks, S., Wake, D. and Shubin, N. 2005. Limb chondrogenesis of the seepage salamander (Desmognathus aenus(Amphibia: Plethodontidae). Journal of Morphology 265:87-101
Read full post
Welcome to The Lancelet!
Add one more biology blog to the list, here comes The Lancelet! Here you'll find the random musings and deluded ramblings of a palaeontology graduate student.
Why "The Lancelet"? This is the common name for an animal sometimes known as Amphioxus (Branchiostoma) - a primitive chordate that has long functioned as a living archetype for the ancestor of all vertebrates... maybe that will be a post for later this week: the history of The Lancelet... but I'm sure there are a number of those on the web.
Books I'm reading:
The Vertebrate Body by A.S. Romer and T. Parsons. Because I never received any formal training in anatomy while I was an undergraduate, I now have to spend my time teaching these important lessons to myself.
From DNA to Diversity S. Carroll et al. Because, when one reads about anatomy they want to know "how the hell did this evolve!?" Carroll and co-authors have bring the complex answers to this question down to lowly wannabe anatomist like me. I recommend it for anyone with a working knowledge of molecular biology.
In the meantime, I'll try to get some news up that the other science bloggers aren't already crawling all over.
Read full post
Why "The Lancelet"? This is the common name for an animal sometimes known as Amphioxus (Branchiostoma) - a primitive chordate that has long functioned as a living archetype for the ancestor of all vertebrates... maybe that will be a post for later this week: the history of The Lancelet... but I'm sure there are a number of those on the web.
Books I'm reading:
The Vertebrate Body by A.S. Romer and T. Parsons. Because I never received any formal training in anatomy while I was an undergraduate, I now have to spend my time teaching these important lessons to myself.
From DNA to Diversity S. Carroll et al. Because, when one reads about anatomy they want to know "how the hell did this evolve!?" Carroll and co-authors have bring the complex answers to this question down to lowly wannabe anatomist like me. I recommend it for anyone with a working knowledge of molecular biology.
In the meantime, I'll try to get some news up that the other science bloggers aren't already crawling all over.
Read full post
Subscribe to:
Posts (Atom)